Evolution and the Problem of Others Minds – Jan 24
Elliott Sober*
Department of Philosophy
University of Wisconsin, Madison 53706
ersober@facstaff.wisc.edu
I.
The following diagram illustrates two inference problems. First, there
is the strictly third-person behavior-to-mind problem in which I observe
your behavior and infer that you occupy some mental state. Second, there
is the self-to-other problem, in which I notice that I always or usually
occupy some mental state when I behave in a particular way; then, when
I observe you produce the same behavior, I infer that you occupy the same
mental state. This second inference is the subject of the traditional philosophical
problem of other minds.
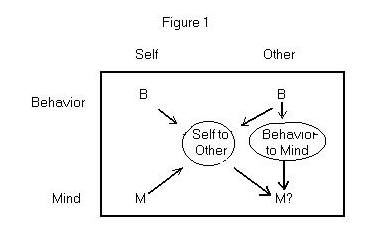
How are these problems related? Notice that the inputs to the behavior-to-mind
problem are a subset of the inputs to the self-to-other problem. In the
first, I consider the behavior of the other individual; in the second,
I consider that behavior, as well as my own behavior and mental state.
This suggests that if the self-to-other problem is insoluble because the
evidence available is too meager, the same will be true of the behavior-to-mind
problem as well. Formulated in this way, it makes no sense to think that
inferences from self to other cannot be drawn, but that purely third-person
behavior-to-mind inferences are sound.
I see no reason to think that a purely third-person scientific psychology
is impossible. Of course, if one is a skeptic about all nondeductive
inference, that skepticism will infect the subject matter of psychology.
And if one thinks that scientific inference can never discriminate
between empirically equivalent theories, one also will hold that science
is incapable of discriminating between such theories when their subject
matter is psychological. However, these aren’t special problems
about psychology. What, then, becomes of the problem of other minds? If
that problem concerns the tenability of self-to-other inference, it appears
to be no problem, if science is able to carry out behavior-to-mind
inferences.
To see how the self-to-other problem can be detached from the problem
of strictly
third-person behavior-to-mind inference, we need to distinguish absolute
from incremental versions of the self-to-other problem. The absolute
problem concerns whether certain input information permits me to infer
that the other person occupies mental state M rather than some alternative
state A. As I’ve said, if information about the behavior of others permits
me to infer that they are in mental state M, then it is hard to see why
this inference should be undermined by adding the premiss that I
myself am in mental state M. However, the question remains of whether first-person
information makes a difference. To whatever degree third-person
information provides an indication of whether the other person has M or
A, does the addition of first-person information modify this assessment?
The incremental version of the self-to-other problem is neutral on the
question of whether third-person behavior-to-mind inference is possible.
It is this incremental problem that I think forms the core of the problem
of other minds, and this is the problem that I want to address here.
Although the problem of other minds usually begins with an introspective
grasp of one’s own mental state, it can be detached from that setting and
formulated more generally as a problem about “extrapolation.” Thus, we
might begin with the assumption that human beings produce certain behaviors
because they occupy particular mental states and ask whether this licenses
the conclusion that members of other species that exhibit the behavior
do so for the same reason. However, none of us knows just by introspection
that all human beings who produce a given behavior do so because
they occupy some particular mental state. In fact, this formulation of
the problem of other minds, in which it is detached from the concept of
introspection, is usually what leads philosophers to conclude that inferences
about other minds from one’s own case are extremely weak. The fact that
I own a purple bow tie isn’t evidence that you do too. This point about
bow ties is supposed to show that the fact that I have a mind isn’t evidence
that you do. I know that I own a purple bow tie, but not by introspection.
Discussion of the problem of other minds in philosophy seems to have
died down (if not out) around thirty years ago. Before then, it was discussed
as an instance of “analogical” or “inductive” reasoning and the standard
objection was that an inference about others based on your own situation
is an extrapolation from too small a sample. This problem does not disappear
merely by thinking of introspective experience as furnishing you with thousands
of data points. The fact remains that they all were drawn from the same
urn -- your own mind. How can sampling from one urn help you infer the
composition of another?
When the problem is formulated in this way, it becomes pretty clear
that what is needed is some basic guidance about inductive inference. It
isn’t the mental content of self-to-other inference that makes it
problematic, but the fact that it involves extrapolation. Some extrapolations
make sense and other don’t. It seems sensible to say that thirst makes
other people drink water, based on the fact that this is usually why I
drink. Yet, it seems silly to say that other folks walk down State Street
at lunchtime because they crave spicy food, based just on the fact that
this is what sets me strolling. By the same token, it seems sensible to
attribute belly buttons to others, based on my own navel gazing. Yet, it
seems silly to universalize the fact that I happen to own a purple bow
tie. What gives?
The first step towards answering this question started to emerge in
the 1960's, not in philosophy of mind, but in philosophy of science. There
is a general point about confirmation that we need to take to heart --
observations provide evidence for or against a hypothesis only in the context
of a set of background assumptions. If the observations don’t deductively
entail that the hypothesis of interest is true, or that it is false, then
there is no saying whether the observations confirm or disconfirm, until
further assumptions are put on the table. This may sound like the Duhem/Quine
thesis, but that way of thinking about the present point is somewhat misleading,
since Duhem (1914) and Quine (1953) discussed deductive, not probabilistic,
connections of hypotheses to observations. If we want a person to pin this
thesis to, it should be I.J. Good. Good (1967, 1968) made this point forcefully
in connection with Hempel’s (1965, 1967) formulation of the ravens paradox.
Hempel thought it was clear that black ravens and white shoes both
confirm the generalization that all ravens are black. The question that
interested him was why one should think that black ravens provide stronger
confirmation than white shoes. Good responded by showing that empirical
background knowledge can have the consequence that black ravens actually
disconfirm
the generalization. Hempel replied by granting that one could have special
information that would undercut his assumption that black ravens and white
shoes both confirm. However, he thought that in the absence of such information,
it was a matter of logic that black ravens and white shoes are confirmatory.
For this reason, Hempel asked the reader to indulge in a “methodological
fiction.” We are to imagine that we know nothing at all about the world,
but are able to ascertain of individual objects what colors they have and
whether they are ravens. We then are presented with a black raven and a
white shoe; logic alone is supposed to tell us that these objects are confirming
instances. Good’s response was that a person in that circumstance would
not be able to say anything about the evidential meaning of the observations.
I think that subsequent work on the concept of evidence, both in philosophy
and in statistics, has made it abundantly clear that Good was right and
Hempel was wrong. Confirmation is not a two-place relationship between
observations and a hypothesis; it is a three-place relation between observations,
a hypothesis, and a background theory. Black ravens confirm the generalization
that all ravens are black, given some background assumptions, but fail
to do so, given others. And if no background assumptions can be brought
to bear, the only thing one can say is -- out of nothing, nothing comes.
If we apply this lesson to the problem of other minds, we obtain the
following result:
The fact that I usually or always have mental property M when I perform
behavior B is, by itself, no reason at all to think that you usually
or always have M when you perform B. If that sounds like skepticism, so
be it. However, the inference from Self to Other can make sense, once additional
background assumptions are stated. A nonskeptical solution to the problem
of other minds, therefore, must identify plausible further assumptions
that bridge the inferential gap between Self and Other.
II.
The problem of other minds has been and continues to be important in
psychology (actually, in comparative psychology), except that there
it is formulated in the first person plural. Suppose that when human beings
perform behavior B, we usually or always do so because we have mental property
M. When we observe behavior B in another species, should we take the human
case to count as evidence that this species also has mental property M?
For this problem to be nontrivial, we assume that there is at least one
alternative internal mechanism, A, which also could lead organisms to produce
the behavior. Is the fact that humans have M evidence that this other species
has M rather than A?
Discussion of this problem in comparative psychology has long been dominated
by the fear of naive anthropomorphism (Dennett 1989, Allen and Bekoff 1997).
C. Lloyd Morgan (1903) suggested that if we can explain a nonhuman organism’s
behavior in terms of a “higher” mental faculty, or in terms of a “lower”
mental faculty, that we should prefer the latter explanation. Morgan’s
successors generally liked Morgan’s canon because of its prophylactic qualities
-- it reduces the chance of a certain type of error. However, it is important
to recognize that there are two types of error that might occur
in this situation:
|
O lacks M |
O has M |
| Deny that O has M |
|
type-2 error |
| Affirm that O has M |
type-1 error |
|
Morgan’s canon does reduce the chance of type-1 error, but that isn’t
enough to justify the canon. By the same token, a principle that encourages
anthropomorphism would reduce the chance of type-2 error, but that wouldn’t
justify this liberal principle, either.
Morgan tried to give a deeper defense of his canon. He thought he could
justify it on the basis of Darwin’s theory of evolution. I have argued
elsewhere that Morgan’s argument does not work (Sober 1998b). However,
it is noteworthy that Morgan explicitly rejects what has become a fairly
standard view of his principle -- that it is a version of the principle
of parsimony. Morgan thought that the simplest solution would be
to view nonhuman organisms as just like us. The canon, he thought, counteracts
this argument from parsimony. As we will see, Morgan’s view about the relationship
of his canon to the principle of parsimony was prescient.
I now want to leave Morgan’s late-nineteenth century ideas about comparative
psychology behind, and fast forward to the cladistic revolution in evolutionary
biology that occurred in the 1970's and after. The point of interest here
is the use of a principle of phylogenetic parsimony to infer phylogenetic
relationships among species, based on data concerning their similarities
and differences (see, for example, Eldredge and Cracraft 1980). Although
philosophers often say that parsimony is an ill-defined concept, its meaning
in the context of the problem of phylogenetic inference is pretty clear.
The hypotheses under consideration all specify phylogenetic trees. The
most parsimonious tree is the one that requires the smallest number of
changes in character state in its interior to produce the observed distribution
of characteristics across species at the tree’s tips.
Consider the problem of inferring how sparrows, robins, and crocodiles
are related to each other. Two hypotheses that might be considered are
depicted in Figure 2. The (SR)C hypothesis says that sparrows and robins
have a common ancestor that is not an ancestor of crocs; the S(RC) hypothesis
says that it is robins and crocs that are more closely related to each
other than either is to sparrows. Now consider an observation -- sparrows
and robins have wings, while crocodiles do not. Which phylogenetic hypothesis
is better supported by this observation?
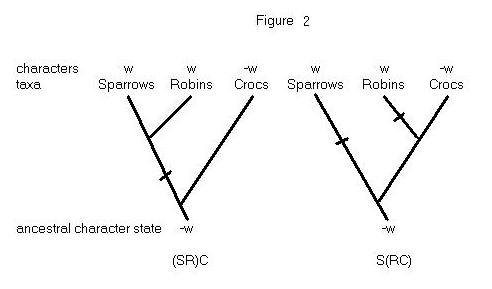
If winglessness is the ancestral condition, then (SR)C is the more parsimonious
hypothesis. This hypothesis can explain the data about tip taxa by postulating
a single change in character state on the branch with a slash through it.
The S(RC) hypothesis, on the other hand, must postulate at least two changes
in character states to explain the data. The principle of cladistic parsimony
says that the observations favor (SR)C over S(RC). Notice that the parsimoniousness
of a hypothesis is assessed not by seeing how many changes it says actually
occurred, but by seeing what the minimum number of changes is that
the hypothesis requires. (SR)C is the more parsimonious hypothesis because
it entails a lower minimum.
Although cladistic parsimony first attracted the attention of biologists
because it helps one reconstruct genealogical relationships, there is a
second type of problem that parsimony is used to address. If you know what
the genealogy is that connects a set of contemporaneous species, and you
observe what character states these tip species have, you can use parsimony
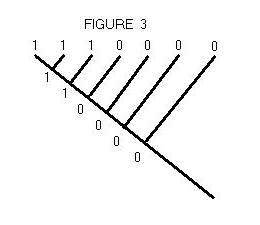
to reconstruct the character states of ancestors. This inference is illustrated
in Figure 3. Given the character states (1's and 0's) of tip species, the
most parsimonious assignment of character states to interior nodes is the
one shown.
Parsimony is now regarded in evolutionary biology as a reasonable way
to infer phylogenetic relationships. Whether it is the absolutely best
method to use, in all circumstances, is rather more controversial. And
the foundational assumptions that need to be in place for parsimony to
make sense as an inferential criterion are also a matter of continuing
investigation.
The reason I’ve explained the basic idea behind cladistic parsimony
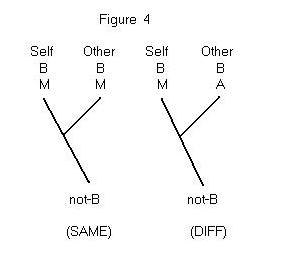
is that it applies to the problem of other minds, when Self and Other are
genealogically related. Suppose both Self and Other are known to have behavioral
characteristic B, and that Self is known to have mental characteristic
M. The question is whether Other should be assigned M as well. As before,
I’ll assume that M is sufficient for B, but not necessary (an alternative
internal mechanism, A, also could produce B). The two hypotheses we need
to consider are depicted in Figure 4. If the root of the tree has the characteristic
not-B ( that is, the trait of having neither-M-nor-A), then the (Same)
hypothesis is more parsimonious than the (Diff) hypothesis. It is consistent
with (Same) that the postulated similarity between Self and Other is a
homology;
it is possible that the most recent common ancestor of Self and Other had
M, and that M was transmitted unchanged from this ancestor to the two descendants.
The (Same) hypothesis, therefore, requires only a single change in character
state, from neither-M-nor-A to M. In contrast, the (Diff) hypothesis requires
at least two changes in character state.
De Waal (1991, 1999) presents this cladistic argument in defense of
the idea that parsimony favors anthropomorphism -- we should prefer
the hypothesis that other species have the same mental characteristics
that we have when they exhibit the same behavior (see also Sober 1998b
and Shapiro forthcoming). This parsimony argument goes against Morgan’s
canon, just as Morgan foresaw. De Waal adds the reasonable proviso that
the parsimony inference is strongest when Self and Other are closely related.
There is another proviso that needs to be added to this analysis, which
is illustrated in Figure 5. As before, Self and Other are observed to have
behavioral trait B. We know by assumption that Self has the mental trait
M. And the question, as before, is what we should infer about Other --
does it have M or A? The new wrinkle is that there are additional species
depicted in the tree, ones that are known to lack B. The genealogical relationships
that connect these further species to Self and Other entail that the most
parsimonious hypothesis is that trait B evolved twice. It now makes no
difference in parsimony whether one thinks that Other has M or A. What
this shows is that parsimony favors anthropomorphism about mentalistic
properties only when the behaviors in question are thought to be homologous.
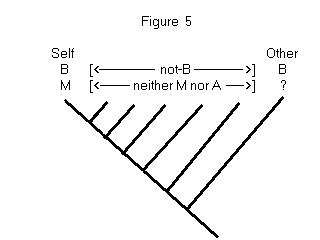
This point has implications about a kind of question that often arises
in connection with sociobiology. Parsimony does not oblige us to think
that “slave-making” in social insects has the same psycho-social causes
as slave-making in humans (or that rape in human beings has the same proximate
mechanism as “rape” in ducks). The behaviors are not homologous,
so there is no argument from parsimony for thinking that the same proximate
mechanisms are at work. This is a point in favor of the parsimony analysis
-- cladistic parsimony explains why certain types of implausible inference
really are implausible. Parsimonious anthropomorphism is not the
same as naive anthropomorphism.
Just as extrapolation from Self to Other can be undermined by behavioral
information about other species, extrapolation also can be undermined by
neurological information about Self and Other themselves. If Self has mental
state M by virtue of being in physical state P1, what are we to make of
the discovery that Other has physical state P2 (where P1 and P2 are mutually
exclusive)? If we accept functionalism’s assurance that mental state M
is multiply realizable, this information does not entail that Other cannot
be in mental state M; after all, P2 might just be a second supervenience
base for M. On the other hand, P2 might be a supervenience base for alternative
state A, and not for M at all. This inference problem is illustrated in
Figure 6. For simplicity, let’s suppose that P1 and P2 each suffice for
B. P1 produces B by way of mental state M; it isn’t known at the outset
whether P2 produces B by way of mental state M, or by way of alternative
state A. We assume that the ancestor at the root of the tree did not exhibit
the behavior in question, and that this ancestor had trait P0.
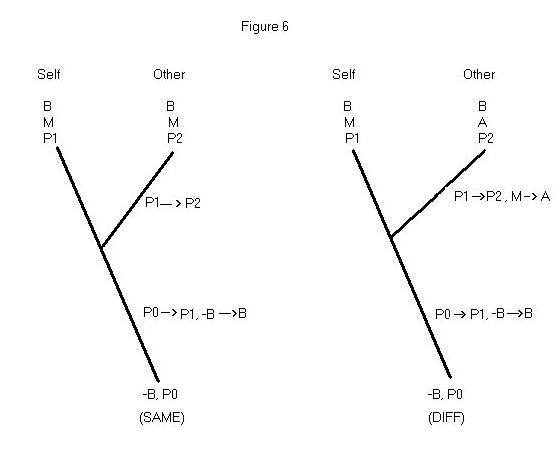
The conclusion we must reach is that the two hypotheses in Figure 6
are equally parsimonious. (Same) construes M as a homology shared by Self
and Other, and P1 and P2 as two of its alternative supervenience bases.
(Same) requires two changes to account for the characteristics at the tips;
either P1 or P2 evolved long ago from an ancestor who had P0, and then
either P1 changed to P2, or P2 changed to P1. Notice that the fact that
not-B changes to B in the same branch in which P0 changes to either P1
or P2 does not count as two changes, since the first entails the
second. The (Diff) hypothesis also requires two changes. The figure depicts
these changes as occurring from P0 to P1 and then from P1 to P2. Of course,
(Diff) also describes a change from M to A on the branch leading to Other,
but this doesn’t count as a change that is distinct from the one on that
branch that goes from P1 to P2. Thus, the discovery of relevant neurophysiological
differences can throw a monkey wrench into the machinery of parsimonious
anthropomorphism.
As a final exercise in understanding how the machinery of phylogenetic
parsimony applies to the problem of extrapolating from Self to Other, let’s
consider the current controversy in cognitive ethology concerning whether
chimps have a theory of mind (see the papers collected in Carruthers and
Smith 1995 and the critical review by Heyes 1998). It is assumed at the
outset that chimps have what Dennett (1989) terms first-order intentionality;
they are able to formulate beliefs and desires about the extra-mental objects
in their environment. The question under investigation is whether they,
in addition, have second-order intentionality. Do they have the ability
to formulate beliefs and desires about the mental states of Self and Other?
Adult human beings have both first- and second-order intentionality. Do
chimps have both, or do they have first-order intentionality only?
Figure 7 provides one cladistic representation that this question might
be given. I assume that the ancestral condition is the absence of both
types of intentionality. The point to notice is that parsimony considerations
do not discriminate between the two hypotheses. The (SAME) hypothesis and
the (DIFF) hypothesis both require two changes in the tree’s interior –
first- and second-order intentionality must each evolve at least once.
The principle of parsimony in this instance tells us to be agnostic, and
so disagrees with Morgan’s canon, which tells us to prefer (DIFF) (Sober
1998b).
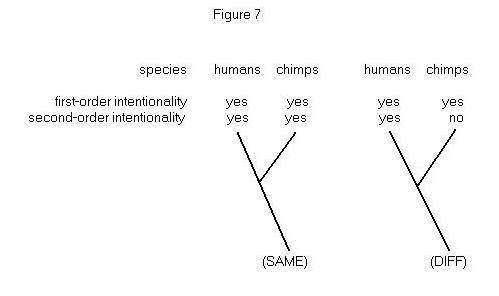
Why does the problem formulated in Figure 7 lead to a different conclusion
from the problem represented in Figure 4 in terms of the two states M and
A? In the latter, we have a parsimony argument that favors extrapolation;
in the former, we find that extrapolating is no more and no less parsimonious
than not extrapolating. Notice that Figure 4 mentions the behavior B, which
internal mechanisms M and A are each able to produce. Figure 7, however,
makes no mention of the behaviors that first- and second-order intentionality
underwrite. This is the key to understanding why the analyses come out
differently.
The point behind Figure 4 is this: if two species exhibit a homologous
behavior, each of them must also possess an internal mechanism (M or A)
for producing the behavior. It is more parsimonious to attribute the same
mechanism to both Self and Other than to attribute different mechanisms
to each. The situation depicted in Figure 7 is different. First-order intentionality
presumably allows organisms that have it to exhibit a range of behaviors
B1. When a species that has first-order intentionality evolves second-order
intentionality as well, this presumably augments the behaviors that organisms
in the species are able to produce; the repertoire expands to B1&B2.
If human beings exhibit B1 because they have first-order intentionality,
we parsimoniously explain the fact that chimps also exhibit B1 (if they
do) by saying that chimps have first-order intentionality. However, there
is no additional gain in parsimony to be had from attributing second-order
intentionality to them as well, unless we also observe them producing the
behaviors in B2. This is why cognitive ethologists are entirely correct
in thinking that they need to identify a behavior that chimps will produce
if they have second-order intentionality, but will not produce if they
possess first-order intentionality only. Identifying a behavior of this
sort permits an empirical test to be run concerning whether chimps have
a theory of mind. Phylogenetic parsimony does not tell us what to expect
here. This example illustrates another respect in which parsimonious anthropomorphism
differs from naive anthropomorphism. Cladistic parsimony does not provide
a blanket endorsement of extrapolation; rather, the principle says that
one should extrapolate when and only when this increases the parsimony
of the total phylogenetic picture.
Although the representation of the problem in Figure 7 is flawed because
it does not indicate which behaviors the two species are able to produce,
it does have the virtue of coding the mentalistic traits more perspicuously.
We should not code the possession of both first- and second-order intentionality
as “M” and the state of having first- but not second-order intentionality
as “A.” This obscures the fact that M is really two traits and that M and
A have a common element. The fact that a cladistic analysis is sensitive
to how the data are coded is not a special consequence of applying parsimony
considerations to traits that are mentalistic; it is a universal feature
of the methodology.
III.
Why should we trust the parsimony arguments just described? Is parsimony
an inferential end in itself or is there some deeper justification for
taking parsimony seriously? In the present circumstance at least, there
is no need to regard parsimony as something that we seek for its own sake.
My suggestion is that parsimony matters in problems of phylogenetic inference
only to the extent that it reflects likelihood (Sober 1988). Here
I am using the term “likelihood” in the technical sense introduced by R.A.
Fisher. The likelihood of a hypothesis is the probability it confers on
the observations, not the probability that the observations confer on the
hypothesis. The likelihood of H, relative to the data, is Pr(Data ˝
H), not Pr(H ˝ Data).
To see how the likelihood concept can be brought to bear on cladistic
parsimony, let’s consider Figure 2. It can be shown, given some minimal
assumptions about the evolutionary process, that the data depicted in that
figure are made more probable by the (RS)C hypothesis than they are by
the R(SC) hypothesis (Sober 1988). These assumptions are as follows:
-- heritability: Character states of ancestors and descendants
are positively correlated. That is, Pr(Descendant has a wing ˝
Ancestor has a wing) > Pr(Descendant has a wing ˝
Ancestor lacks a wing).
-- chance: All probabilities are strictly between 0 and 1.
-- screening-off: Lineages evolve independently of each other,
once they branch off from their most recent common ancestor.
I think that most biologists would agree that these assumptions hold
pretty generally. The first of them does not say that descendants
probably end up resembling their ancestors. The claim is not that
stasis is more probable than change -- that Pr(Descendant has a wing ˝
Ancestor has a wing) > Pr(Descendant lacks a wing ˝
Ancestor has a wing). Rather, the claim is that if a descendant has a wing,
that this result would have been more probable if its ancestor had had
a wing than it would have been if its ancestor had lacked a wing. This
is equivalent to saying that the trait in question has nonzero heritability.
Perhaps the only assumption open to serious question is the last one --
after all, there are ecological circumstances in which a lineage’s evolving
a trait influences the probability that other contemporaneous lineages
will do the same. But even here, the assumption of perfect independence
could be weakened without materially affecting the qualitative conclusions
I want to draw. And in many circumstances, this assumption is a reasonable
idealization. What we have here is a result that Reichenbach (1956) proved
in connection with his principle of the common cause (Sober 1988)
.
Not only does likelihood provide a framework for understanding the role
of parsimony considerations in phylogenetic inference; it also allows us
to address the Self and Other problem depicted in Figure 4. The assumptions
listed above entail the following inequality:
(P) Pr(Self has M ˝ Other has M) > Pr(Self
has M ˝ Other has A).
Provided that Self and Other are genealogically related, there is a
likelihood justification for anthropomorphism. The observation that Self
has M is rendered more probable by the hypothesis that Other has M than
by the hypothesis that Other has A. Such differences in likelihood are
generally taken to indicate a difference in support -- the observation
favors the first hypothesis over the second (Royall 1997, Sober 1999).
Turning to the concept of heritability, let’s consider the factors that
influence the values of conditional probabilities that have the form Pr(Descendant
has ±M ˝ Ancestor has ±M). One natural way to
understand such probabilities, whether they apply to individual organisms
or to species, is to think of the lineage linking ancestor to descendant
as having a given, small probability of changing from M to A in any brief
moment of time, and another, possibly different, small probability of changing
in the opposite direction in that same small instant of time. Then the
probability that a lineage ends in one state, given that it begins in another,
will be a function of those “instantaneous” probabilities of change and
of the amount of time that there is in the lineage. The standard analysis
of these lineage transition probabilities entails that Pr(Descendant has
M ˝ Ancestor has M) is close to 1 when the amount
of time is very small and that
Pr(Descendant has M ˝ Ancestor has M) = Pr(Descendant
has M ˝ Ancestor has A)
when the amount of time separating ancestor and descendant is infinite
(Sober 1988). It follows that the inequality stated in (P) is maximal when
Self and Other are very closely related and becomes smaller as Self and
Other become more distantly related. This model confirms what De Waal suggests
-- that anthropomorphism is on firmer ground for our near relatives than
it is for those species to whom we are related more distantly. This point
need not be added as an independent constraint on parsimony arguments (which
have no machinery for taking account of recency of divergence), but flows
from the likelihood analysis itself.
A probability framework also throws light on the ideas discussed in
connection with Figure 5. In this case there is no difference in parsimony
between assigning M to Other and assigning A to Other, if the behavior
B shared by Self and Other is a result of convergence.
What probabilistic assumptions might underwrite this conclusion? The
assumptions just discussed that suffice to justify proposition (P) are
not enough. What is needed, in addition, is some way to compare the probability
of M’s arising and of A’s arising in lineages that have neither. If these
probabilities are the same, then the likelihoods of the two hypotheses
will be the same. And if no assumption about how these probabilities are
related can be endorsed, then
no extrapolation can be endorsed. Likelihood considerations do not oblige
us to think that slave-making in the social insects is driven by the same
internal mechanisms that lead human beings to enslave each other.
IV.
Genealogical relatedness suffices to justify a likelihood inference
from Self to Other, if the Reichenbachian assumptions that I described
hold true. However, is genealogical relatedness necessary for this
extrapolation to make sense? Proposition (P) says that Self and Other are
correlated with respect to trait M. What could induce this correlation?
Reichenbach argued that whenever there is a correlation of two events,
either the one causes the other, or the other causes the one, or the two
trace back to a common cause. Considerations from quantum mechanics suggest
that this isn’t always the case (Van Fraassen 1982a), and doubts about
Reichenbach’s principle can arise from a purely classical point of view
as well (Van Fraassen 1982b; Sober 1988; Cartwright 1989). However, if
we are not prepared to suppose that mentalistic correlations between Self
and Other are brute facts, and if my having M does not causally influence
whether Other has M (or vice versa), then Reichenbach’s conclusion
seems reasonable, if not apodictic – if Self and Other are correlated,
this should be understood as arising from a common cause.
Genealogical relatedness is one type of common cause structure. It can
induce the correlation described in (P) by having ancestors transmit genes
to their descendants, but there are
alternatives that we need to recognize. To begin with, parents exert
nongenetic
influences on their offspring through teaching and learning. For example,
children have a higher probability of speaking Korean if their parents
speak Korean than if their parents do not, but this is not because there
is a gene for speaking Korean. And there are nongenetic connections between
parents and offspring that do not involve learning, as when a mother transmits
immunity to her children through her breast milk.
Correlations between Self and Other also can be induced by common causes
when Self and Other are not genealogically related. If students resemble
their teachers, then students of the same teacher will resemble each other.
Here learning does the work that genetic transmission is also able to do.
Similarly, Self and Other can be correlated when they are influenced by
a common environmental cause that requires no learning. For example, if
influenza is spreading through one community, but not through another,
then the fact that I have the flu can be evidence that Other does too,
if the two of us live in the same community.
I list these alternatives, not because all of them apply with equal
plausibility to the problem of other minds, but to give an indication of
the range of alternatives that needs to be considered. The question is
whether there are common causes that impinge on Self and Other that induce
the correlation described in (P); if there are, then there will be a likelihood
justification for extrapolating from Self to Other.
V.
What, exactly, does cladistic parsimony and its likelihood analysis
tell us about the problem of other minds? When I cry out, wince, and remove
my body from an object inflicting tissue damage, this is (usually) because
I am experiencing pain. Is the same set of behaviors, when produced by
other organisms, whether they are human or not, evidence that they feel
pain? This hypothesis about Other is more parsimonious, and it is
more likely, if Reichenbachian assumptions about common causes are correct.
Does that completely solve the problem of other minds, or does there remain
a residue of puzzlement?
One thing that is missing from this analysis is an answer to the question
-- how much evidence does the introspected state of Self provide
about the conjectured state of Other? I have noted that the likelihoods
in proposition (P) become more different as Self and Other become more
closely related, but this comparative remark does not entail any quantitative
benchmarks. It is left open whether the observations strongly favor
one hypothesis over the other, or do so only weakly.
Another detail that I have not addressed is how probable it is
that Other has M. If I have M and Self and Other both exhibit behavior
B, is the probability greater than 0.5 that Other has M as well? The discussion
here helps answer that question, in that principle (P) is equivalent
to the claim that Self’s having M raises the probability that Other has
M. Whatever the prior probability is that Other has M, the posterior probability
is greater. Whether additional information can be provided that allows
the value of that posterior probability to be estimated is a separate question.
A genealogical perspective on the problem of other minds helps clarify
how that problem differs from the behavior-to-mind problem discussed at
the outset. At first glance, it might appear that it doesn’t matter to
the problem of other minds whether the other individual considered is a
human being, a dog, a Martian, or a computer. In all these cases, the question
can be posed as to whether knowledge of one’s own case permits extrapolation
to another system that is behaving similarly. We have seen that these different
formulations receive different answers. I share ancestors with other human
beings, and I share other, more remote, ancestors with nonhuman organisms
found on earth. However, if creatures on other planets evolved independently
of life on earth, then I share no common ancestors with them. In this case,
extrapolation from Self to Other does not have the justification I have
described. I would go further and speculate that it has no justification
at all. This doesn’t mean that we are never entitled to attribute mental
states to such creatures. What it does mean is that we must approach such
questions as instances of the purely third-person behavior-to-mind inference
problem. Similar remarks apply to computers. When they behave similarly
to us (perhaps by passing an appropriate Turing test), we may ask what
causes them to do so. We have no ancestors in common with them; rather,
we have constructed them so that they produce certain behaviors. However,
this provides no reason to think that the proximate mechanisms behind those
behaviors resemble those found in human beings. For both Martians and computers,
extrapolation from one’s own case will not be justified. Indeed, this point
applies to organisms with whom we do share ancestors, if the behaviors
we have in common with them are not homologies. And even when Self and
Other do share a behavioral homology, if Self and Other are known to deploy
different neural machinery for exhibiting that behavior, the extrapolation
of M from Self to Other is undermined.
A probabilistic representation of the problem of other minds shows that
the usual objection to extrapolating from Self to Other is in fact irrelevant,
or, more charitably, that it rests on a factual assumption about the world
that we have no reason to believe. The question is not whether introspected
information about one’s own mind provides lots of data or only a little.
Rather, the issue is how strong the correlation is between Self and Other.
Consider two urns that are filled with balls; each ball has one color,
but the frequencies of different colors in the urns are unknown. The urns
may be similar or identical in their compositions, or they may be very
different. If I sample one ball from the first urn and find that it is
green, does this provide much information for making a claim about the
second urn’s composition? If I sample a thousand balls from the first urn,
does this allow me to say any more about the second urn? Everything depends
on how the two urns are related. If they are independent, then samples
drawn from the first, whether they are small or large, provide no information
about the second. But if they are not independent, then even a small sample
from the first may be informative with respect to the second.
There is nothing wrong with asking whether my introspected knowledge
of Self
extrapolates to a well-supported conclusion about Other. But the skeptical
assertion
that it does not involves a factual claim about the world. Claims of independence
are no more a priori than are claims of correlation. Perhaps a special
creationist would want to assume that different species are uncorrelated,
but no evolutionist would think that this is so. And creationists and evolutionists
agree that all human beings are genealogically related, so here
it is especially problematic to assume that Self and Other are independent.
The argument I have presented is intended to show how certain propositions
are justified; I have not addressed the question of whether people
are justified in believing this or that proposition. My own mental state
can be an indicator of the mental states of others, whether or not I know
that this is true, or understand why it is true. But what is required
for people to be justified in extrapolating from Self to Other? Must they
have reason to believe that principle (P) is satisfied? Or is it enough
that (P) is satisfied? I will not enter into this epistemological
thicket concerning internalist versus externalist views of justification,
except to note that the ideas surveyed here are relevant across a range
of views that one might take. For the externalist, it is enough that a
common cause structure has induced the correlation described by proposition
(P). For the internalist, the point of interest is that individuals are
entitled to extrapolate from Self to Other if they already have the justified
belief that a common cause structure has rendered (P) true.
Is it possible for me to figure out that proposition (P) is true, if
I don’t already
know whether Other has M or A? Surely I can. I can tell whether M is
heritable by looking at still other individuals who are known to have either
M or A and see how they are genealogically related. But is it possible
for me to determine that (P) is true without knowing anything about which
individuals (other than myself) have M and which have A? Without this information,
how can I determine whether the traits are heritable? To be sure, I can
tell whether the behavior B is heritable, but what of the two possible
internal mechanisms that can cause individuals to exhibit B? Well, knowledge
in some strong philosophical sense is probably unnecessary, but perhaps
judgments about heritability require one to have reasonable opinions, however
tentative, about which individuals have which traits. Even if that were
so, the solution to the problem of other minds that I have suggested would
not be undermined. The incremental version of the problem, recall, asks
whether knowledge of my own case makes a difference in the characteristics
I attribute to others. It isn’t required that I conceive of myself as beginning
with no knowledge at all concerning the internal states of others.
We learned from I.J. Good that there is no saying whether a black raven
confirms the generalization that all ravens are black unless one is prepared
to make some substantive background assumptions. The mere observation that
the object before you is a black raven is not enough. The same point, applied
to the problem of other minds, is that the mere observation that Self and
Other share B and that Self has M is not enough. Further assumptions are
needed to say whether these observations confirm the hypothesis that Other
has M. Recognizing this point in the ravens paradox does not lead inevitably
to skepticism, and it should not have that effect in the case of self-to-other
inference. The problem of other minds should not be shackled with the “methodological
fiction” that Hempel imposed on the ravens paradox. When the fetters are
broken, the problem of other minds turns into the problem of searching
out common causes.
References
Allen, C. and Bekoff, M. (1997): Species of Mind – the Philosophy
and Biology of Cognitive Ethology. Cambridge: MIT Press.
Carruthers, P. and Smith, P. (1995): Theories of Theories of Mind.
Cambridge: Cambridge University Press, pp. 277-292.
Cartwright, N. (1989): Nature’s Capacities and Their Measurement.
Oxford: Oxford University Press.
De Waal, F. (1991): “Complementary Methods and Convergent Evidence in
the Study of Primate Social Cognition.” Behaviour 118: 297-320.
De Waal, F. (1999): “Anthropomorphism and Anthropodenial – Consistency
in our Thinking about Humans and Other Animals.” Philosophical Topics,
forthcoming.
Dennett, D. (1989): “Intentional
Systems in Cognitive Psychology -- the ‘Panglossian Paradigm’ Defended.”
In The Intentional Stance. Cambridge: MIT Press, pp. 237-268.
Duhem, P. (1914): The Aim and Structure of Physical Theory. Princeton:
Princeton University Press. 1954.
Eldredge, N. and Cracraft, J. (1980): Phylogenetic Patterns and the
Evolutionary Process. New York: Columbia University Press.
Enç, B. (1995): “Units of Behavior.” Philosophy of Science 62:
523-542.
Forster, M. and Sober, E.
(1994): “How to Tell When Simpler, More Unified, or Less Ad Hoc
Theories Will Provide More Accurate Predictions." British Journal for
the Philosophy of Science 45: 1-36.
Forster, M. and Sober, E.
(2000) “Why Likelihood?” In M. Taper and S. Lee (ed.), The Evidence
Project, Chicago: University of Chicago Press.
Good, I.J. (1967): “The White
Shoe is a Red Herring.” British Journal for the Philosophy of Science
17: 322.
Good, I.J. (1968): “The White
Shoe Qua Herring is Pink.” British Journal for the Philosophy of Science
19: 156-157.
Hempel, C.G. (1965): “Studies
in the Logic of Confirmation.” in Aspects of Scientific Explanation
and Other Essays. New York: Free Press.
Hempel, C.G. (1967): “The
White Shoe -- No Red Herring.” British Journal for the Philosophy of
Science 18: 239-240.
Heyes, C. M. (1998): ‘Theory of Mind in Nonhuman Primates.’ Behavioral
and Brain Sciences 21, 101-148.
Kyburg, H. (1961): Probability
and the Logic of Rational Belief. Middletown, CT: Wesleyan University
Press.
Morgan, C.L. (1903): An
Introduction to Comparative Psychology (2nd ed.). London: Walter Scott.
Quine, W. (1953): “Two Dogmas of Empiricism.” In From a Logical Point
of View. Cambridge, MA: Harvard University Press. 20-46.
Reichenbach, H. (1956): The Direction of Time. Berkeley: University
of California Press.
Royall, R. (1997): Statistical
Evidence -- a Likelihood Paradigm. Boca Raton: Chapman and Hall/CRC.
Shapiro, L. (1999): “Presence
of Mind.” In V. Hardcastle (ed.), Biology Meets Psychology – Constraints,
Connections, and Conjectures. Cambridge: MIT Press.
Shapiro, L. (forthcoming):
“Adapted Minds.” In J. McIntosh (ed.), Naturalism, Evolution, and Intentionality.
Cambridge: MIT Press.
Sober, E. (1988): Reconstructing
the Past -- Parsimony, Evolution, and Inference. Cambridge: MIT Press.
Sober, E. (1998a ): “Black
Box Inference -- When Should an Intervening Variable be Postulated?” British
Journal for the Philosophy of Science 49: 469-498.
Sober, E. (1998b): “Morgan’s Canon.” In C. Allen and D. Cummins (eds.),
The
Evolution of Mind, New York: Oxford University Press, pp.224-242.
Sober, E. (1999): “Testability.” Proceedings and Addresses of the
American Philosophical Association73:
47-76. Also available at the following URL – http://philosophy.wisc.edu/sober.
Sober, E. and Barrett, M.
(1992): “Conjunctive Forks and Temporally Asymmetric Inference.” Australasian
Journal of Philosophy 70: 1-23.
Van Fraassen, B. (1982a):
“The Charybdis of Realism – Epistemological Implications of Bell’s Inequality.”
Synthese
52: 25-38.
Van Fraassen, B. (1982b) “Rational Belief and the Common Cause Principle,”
In R. McLaughlin
(ed.), What? Where? When? Why? Essays in Honor of Wesley Salmon.
Dordrecht: Reidel, pp. 193-209.
Notes
*. My thanks to Colin Allen,
Martin Barrett, Marc Bekoff, Tom Bontly, James Crow, Frans De Waal, Ellery
Eells, Mehmet Elgin, Berent Enç, Peter Godfrey-Smith, Daniel Hausman, Richard
Lewontin, David Papineau, Larry Shapiro, and
Alan Sidelle for useful comments on earlier drafts. I also benefitted from
discussing this paper at the London School of Economics and Political Science
and at University of Illinois at Chicago.